要点
- 海産性の油脂高生産藻類ナンノクロロプシスで、蓄積する脂質の量と脂肪酸組成を改変する技術を開発
- ナンノクロロプシスのリン欠乏に応答した遺伝子発現制御の仕組みが、種の異なる藻類クラミドモナスと類似することを見出し、その仕組みを活用
- リン欠乏に応答する遺伝子プロモーターを種々の脂質合成遺伝子とセットでナンノクロロプシスに導入することで、様々な種類の有用脂肪酸の生産へ応用することを期待
概要
東京工業大学大学院生命理工学研究科の岩井雅子CREST研究員、太田啓之生命理工学研究科/地球生命研究所教授らは、海産性の油脂高生産藻類として注目されるナンノクロロプシス[用語1]を用い、油脂の蓄積量と油脂中の脂肪酸組成を改変する技術を開発した。
多くの藻類では窒素欠乏時に油脂を蓄積することが知られている。その一方、窒素欠乏条件では藻類の生育が著しく阻害されることから、有用油脂の生産においては、生育しながら油脂を貯める手法の開発が課題となっている。岩井らは、高密度な細胞培養が可能で細胞中に油脂だけを多量に貯める海産性の藻類ナンノクロロプシスで、リンの欠乏条件では窒素欠乏条件と比べ、生育を維持しながら油脂を高蓄積することを発見した。さらに、リン欠乏時に、細胞の膜中のリン脂質を糖脂質に転換する緑藻クラミドモナス[用語2]と同様な仕組みがナンノクロロプシスで働いていることも見出した。そこで緑藻クラミドモナスから取得したリン欠乏応答性の糖脂質合成遺伝子プロモーター[用語3]と油脂合成遺伝子を結合してナンノクロロプシスに導入した結果、脂質の蓄積を増強させるとともに、脂肪酸の組成を改変することに成功した。今後、リン欠乏応答プロモーターと種々の脂質合成遺伝子とをセットで藻類に導入することで、様々な高付加価値の油脂が工業規模で生産できると期待される。
- ※
- この研究は太田教授が科学技術振興機構 戦略的創造研究推進事業(CREST) 「藻類・水圏微生物の機能解明と制御によるバイオエネルギー創成のための基盤技術の創出」研究領域(研究総括:松永是(東京農工大学 学長))における研究課題「植物栄養細胞をモデルとした藻類脂質生産系の戦略的構築」の一環として、東工大大学院生命理工学研究科の下嶋美恵准教授、堀孝一CREST研究員と佐々木結子地球生命研究所特任助教との共同で行った。成果の内容は9月7日発行のスイス科学雑誌「フロンティアズ イン マイクロバイオロジー(Frontiers in Microbiology)」に掲載された。
研究の背景と経緯
多くの藻類は窒素欠乏などの栄養欠乏条件下でトリアシルグリセロール(TAG;油脂[用語4])を蓄積することが知られており、近年、藻類が生産するTAGやTAGに含まれる脂肪酸が化学工業用油脂やバイオ燃料の原材料として注目されている。藻類油脂をこのような原材料として活用するためには、利用目的に合った油脂や有用脂肪酸などを大量に生産させる技術の開発が必要となる。しかしながら、本来光合成膜[用語5]や細胞膜などに存在しない有用脂肪酸や油脂を大量に生産させようとすると、それらが光合成膜などに蓄積し、光合成を阻害するという問題がある。これを回避するには、適切な条件下で有用脂肪酸や油脂を生産させながら、それを細胞内に油滴として高蓄積させることが有効である。しかし、一般的に藻類の油脂生産の条件とされる窒素欠乏条件では、光合成や新規脂肪酸合成が抑制され、細胞がほとんど増殖しないため、このような手法を用いることは難しい。
岩井研究員らは、以前の研究で、緑藻クラミドモナスを増殖させながら光合成の場である葉緑体を維持してTAG蓄積をできる条件を探索し、リン欠乏条件では窒素欠乏条件ほど劇的な細胞増殖抑制は見られないことを見出した。さらにその条件下でのTAG蓄積を遺伝子操作によって強化することに成功した。しかし、クラミドモナスの場合、栄養欠乏時にTAGだけでなくデンプンも蓄積し、また高密度での細胞培養が難しいため、藻類での効率的な油脂生産を目指すためには、より高密度で細胞培養ができ、細胞中に油脂だけを多量に貯めることができるナンノクロロプシスなどの油脂高生産藻での生産技術の開発が期待されていた(図1)。
Clik here to view.
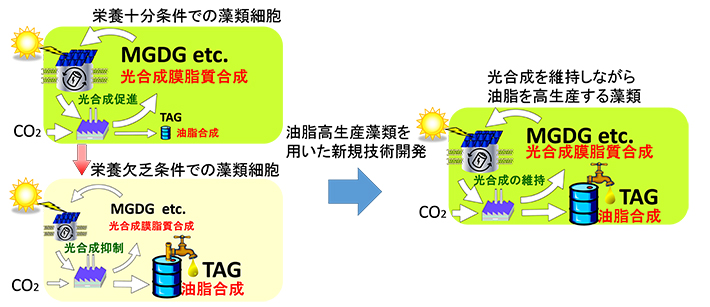
- 図1.
- 藻類の栄養欠乏による油脂の蓄積と油脂高生産藻類を用いた生産技術開発
MGDGは光合成を行う葉緑体に最も主要な膜脂質。窒素欠乏時の藻類では、TAGが蓄積する一方、MGDGなど光合成の膜脂質が減少し、光合成が抑制される。藻類で利用目的に応じた有用脂質を工業規模で生産するためには、油脂の生産性に優れた藻類を活用して、光合成を維持したまま有用な油脂を高生産する技術の開発が必要となる。
研究内容
岩井研究員らはナンノクロロプシスでの窒素欠乏条件とリン欠乏条件の比較(図2)から、ナンノクロロプシスのリン欠乏下では光合成の場である葉緑体を維持したままTAGを蓄積することを見出した。そこでクラミドモナスなどの藻類や植物がリン欠乏に適応する際に起こすリン脂質から糖脂質への膜脂質の転換が、クラミドモナスとは異なり二次共生藻[用語6]に属するナンノクロロプシスでも機能していると予測し、リン欠乏時の膜脂質の変動や膜脂質転換に関わる遺伝子の応答を調べた。その結果、ナンノクロロプシスでもリン欠乏時に糖脂質合成酵素遺伝子の発現が誘導され、リン脂質と糖脂質が置き換わることでリン欠乏に適応することを見出した。
Clik here to view.

- 図2.
- ナンノクロロプシス培養の写真
窒素欠乏条件では細胞の増殖が著しく抑制されているのに対して、リン欠乏では通常条件と同様に緑が濃くなり、細胞が増殖して葉緑体が発達しているのが判る。
図3の通常培養条件に示すように、リンを十分含む条件では葉緑体や細胞の膜脂質(図のMGDGやDGDG、PCなど)が細胞に含まれる脂質の多くを占めるが、リン欠乏条件ではTAGの含量が著しく増大する。また、リン欠乏条件ではTAGの増大と同時に葉緑体の糖脂質の一つであるSQDGが増加しており、別の実験からSQDGの合成にかかわるSQD2遺伝子の発現が顕著に誘導されていることも分かった。そこで、その仕組みを活用して新規脂肪酸を合成させながらTAG蓄積を増強するため、先のクラミドモナスの研究で取得したリン欠乏条件下で発現上昇するクラミドモナスSQDG合成遺伝子(CrSQD2)のプロモーター「pCrSQD2」[用語7]とクラミドモナスのTAG合成酵素「CrDGTT4」の利用を試みた。pCrSQD2とCrDGTT4のセットをナンノクロロプシスに導入したところ、遺伝子を導入した形質転換株では、CrDGTT4遺伝子を導入していない比較対照株(コントロール株)と比べ、リン欠乏条件下でのTAG蓄積がさらに増加した(図3形質転換株参照)。またTAG中の脂肪酸にはCrDGTT4が基質として好むオレイン酸(C18:1(9)=炭素数が18で二重結合を9位の位置に一つ持つ脂肪酸)がコントロール株の約2倍含まれていた(図4)。これらの結果は、二次共生藻として緑藻とは分類上も全く異なる油脂高生産藻ナンノクロロプシスで、緑藻クラミドモナス由来のリン欠乏応答性プロモーターを用いることによりTAG蓄積の増強と脂肪酸組成の改変ができることを示している。
またこの結果は、リン欠乏条件に適応する仕組みそのものが種の異なる緑藻と二次共生藻の間で広く保存されていることを示しており、進化的な観点からも興味深い。
Clik here to view.
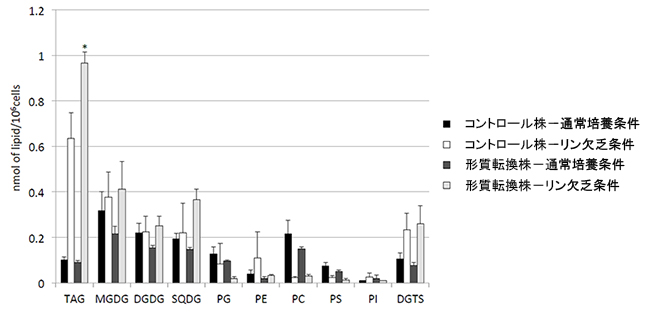
- 図3.
- 細胞中に含まれるTAGと膜脂質の量
全て培養4日目。MGDG、DGDG、SQDGは葉緑体の膜に含まれる糖脂質。PG、PE、PC、PS、PIは葉緑体や細胞の膜に含まれるリン脂質。DGTSは藻類細胞の膜に特有のベタイン脂質。リン欠乏時に細胞中のリン脂質が減少し、TAGやSQDG、DGTSが増加する。特にTAGの増大が顕著で、形質転換株ではコントロール株に比べその増大がさらに強化されているのが判る。
Clik here to view.
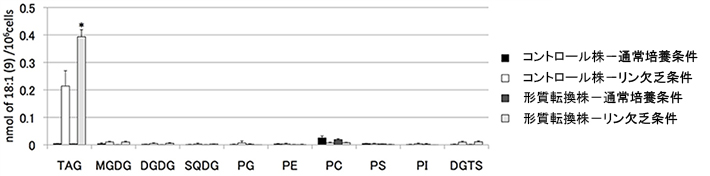
- 図4.
- 形質転換株とコントロール株のそれぞれの脂質に含まれるオレイン酸(C18:1)の量(培養4日目)
オレイン酸は通常培養条件では細胞中にあまり見られないが、リン欠乏時にTAGに蓄積する。形質転換株のオレイン酸はコントロール株の更に2倍に増加した。
今後の展開
本研究により、海産性の油脂高生産藻ナンノクロロプシスで、ナンノクロロプシスと分類上全く種が異なる緑藻クラミドモナスのリン欠乏応答プロモーターを用いてTAG生産の増強と脂肪酸組成の改変ができることが明らかになった。ナンノクロロプシスは他の藻類に比べ、細胞に多量の油脂を蓄積すると同時に、高密度での培養が可能であることが知られており、また海産性藻類であることから海水を用いて培養できる。リン欠乏応答性プロモーターを機能の異なる種々の脂質合成遺伝子と結合して用いることで、ナンノクロロプシスで様々な有用脂肪酸を含む油脂を大量に生産できることが期待される。今回そのプロモーターを利用した糖脂質合成遺伝子SQD2は藻類に広く保存されることが分かっており、他の藻類で同様な手法が活用できることも期待される。
用語説明
[用語1] ナンノクロロプシス : 海産性の油脂高生産藻類。細胞内に細胞の重さの50%を超える脂質(トリアシルグリセロール、TAG)を蓄積することで知られる。このような油を多量に生産する藻類としては他にボトリオコッカスなどがよく知られているが、ボトリオコッカスは、TAGではなく長鎖の炭化水素やテルペンを合成する。長鎖の炭化水素はその性質上ガソリンの代替として期待されているが、ナンノクロロプシスなどが生産するTAGは、軽油の代替としての利用が可能であるといわれており、生産する油によって用途が異なる。
[用語2] クラミドモナス : 緑藻綱クラミドモナス目に属する単細胞藻類。ゲノム解析が進みモデル藻類として用いられる。
[用語3] 栄養(リン)欠乏応答性プロモーター : 栄養(リン)欠乏時に遺伝子の発現が誘導される糖脂質合成酵素遺伝子などの上流域に存在し、下流の遺伝子のリン欠乏時の誘導を制御する働きを持つ。
[用語4] TAG : トリアシルグリセロール。1分子のグリセロールに3分子の脂肪酸がエステル結合した中性脂肪の1つ。藻類や植物の細胞内に油滴として蓄積する。
[用語5] 光合成膜 : 光合成細菌、藻類、植物など、光合成を行う生物の光合成反応をつかさどる高度に発達した細胞内部の膜を示す。特に藻類や植物の細胞に存在する光合成器官である葉緑体は、その内部にチラコイド膜という光合成を担う発達した膜を持つが、このチラコイド膜のことを光合成膜ともいう。
[用語6] 二次共生藻 : 緑藻や高等植物など、原始シアノバクテリアの細胞内共生により光合成器官である葉緑体を獲得した生物を一次共生生物と呼ぶのに対して、緑藻や植物とは全く起源の異なる生物が、葉緑体を持つ他の藻類を細胞内に丸ごと取り込んで光合成の能力を獲得した場合、そのような藻類を二次共生藻と呼ぶ。コンブやワカメも二次共生藻であり、これらの生物は光合成を行うが、緑藻や植物とは全く起源が異なる。
[用語7] クラミドモナスSQD2遺伝子のプロモーター「pCrSQD2」 : SQD2は、藻類や植物の葉緑体に含まれる主要膜脂質スルホキノボシルジアシルグリセロール(SQDG)の合成酵素。SQDGは硫黄を含む糖脂質で、リン欠乏時に特に増加して、葉緑体に含まれるリン脂質であるホスファチジルグリセロール(PG)の代替を担う。クラミドモナスや植物のSQD2遺伝子はリン欠乏時にその発現が強く誘導されるが、遺伝子の上流にあるプロモーター領域がその制御を担う。
論文情報
掲載誌 : |
Frontiers in Microbiology |
論文タイトル : |
Manipulation of oil synthesis in Nannochloropsis strain NIES-2145 with a phosphorus starvation-inducible promoter from Chlamydomonas reinhardtii |
著者 : |
Masako Iwai, Koichi Hori, Yuko Sasaki-Sekimoto, Mie Shimojima and Hiroyuki Ohta |
DOI : |
10.3389/fmicb.2015.00912 Image may be NSFW. Clik here to view.  |
東京工業大学地球生命研究所について
地球生命研究所(ELSI)は、文部科学省が平成24年に公募を実施した世界トップレベル研究拠点プログラム(WPI※)に採択され、同年12月7日に産声をあげた新しい研究所。「地球がどのように出来たのか、生命はいつどこで生まれ、どのように進化して来たのか」という、人類の根源的な謎の解明に挑んでいる。
- ※
- 世界トップレベル研究拠点プログラム(WPI)は、平成19年度から文部科学省の事業として開始されたもので、システム改革の導入などの自主的な取組を促す支援により、第一線の研究者が是非そこで研究したいと世界から多数集まってくるような、優れた研究環境ときわめて高い研究水準を誇る「目に見える研究拠点」の形成を目指している。

問い合わせ先
東京工業大学 大学院生命理工学研究科
教授 太田啓之
Email : ohta.h.ab@m.titech.ac.jp
Tel : 045-924-5736 / Fax : 045-924-5527
東京工業大学 広報センター
Email : media@jim.titech.ac.jp
Tel : 03-5734-2975 / Fax : 03-5734-3661
科学技術振興機構 広報課
Email : jstkoho@jst.go.jp
Tel : 03-5214-8404 / Fax : 03-5214-8432
JST事業に関すること
科学技術振興機構 戦略研究推進部 川口 哲
Email : crest@jst.go.jp
Tel : 03-3512-3524 / Fax : 03-3222-2064